Ik help jou om in een druk bestaan fitter door het leven te gaan
Online personal training
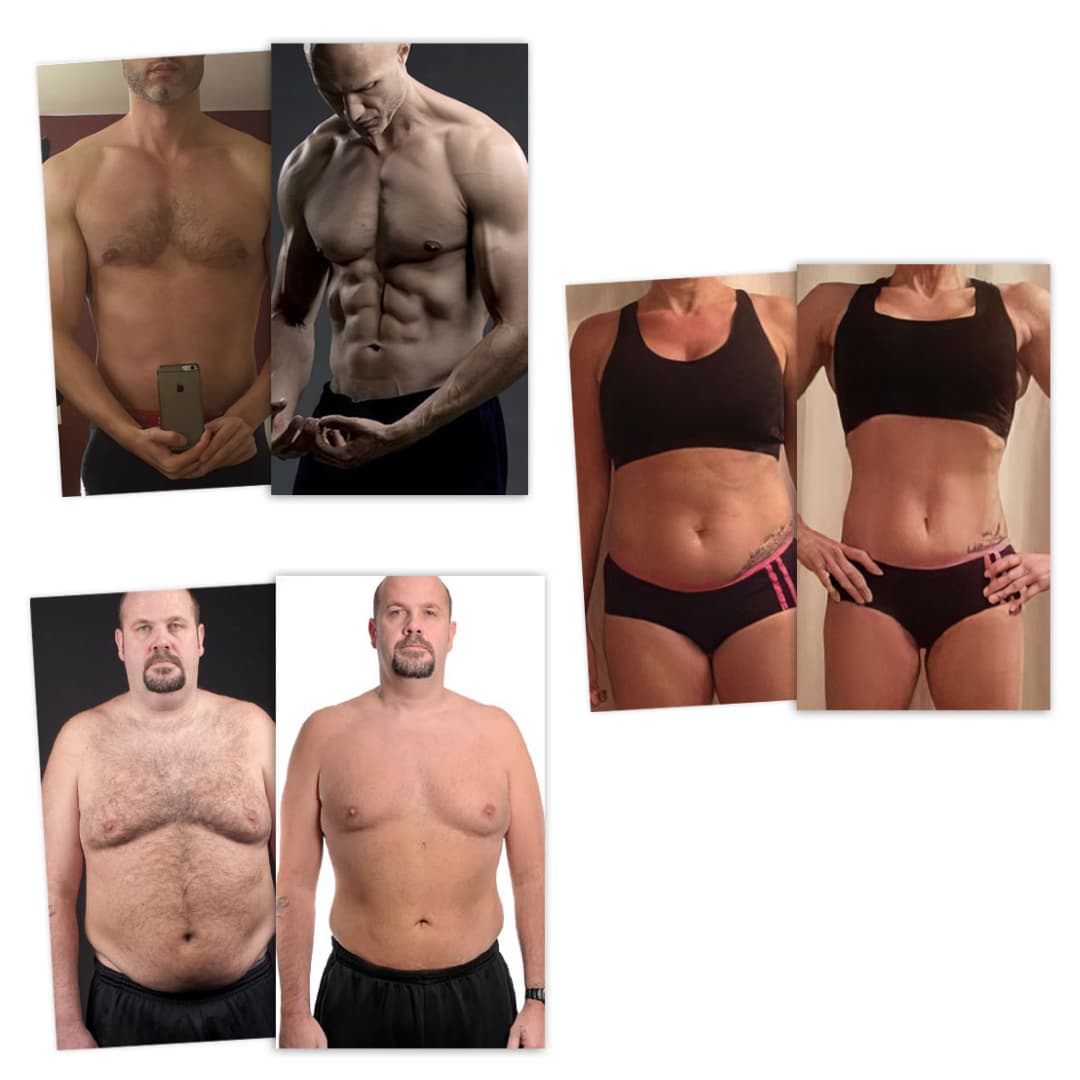
Als online personal trainer help ik jou om in een druk bestaan gespierder, fitter en slanker door het leven te gaan.
Tegenwoordig is er via het internet zoveel informatie te vinden omtrent vetverlies, voeding en spieropbouw, dat je door de bomen het bos niet ziet. Hierdoor is het moeilijk om feiten te onderscheiden van fabels.
Doordat ik de afgelopen jaren veel kennis heb opgedaan op het gebied van vetverlies, spieropbouw en gedragsverandering en honderden klanten heb geholpen, heb ik veel inzicht gekregen van wat werkt.
Ik combineer een wetenschappelijk onderbouwde aanpak met meer dan 10 jaar praktijkervaring in mijn coaching. Via mijn app en videogesprekken help ik jou met jouw fysieke doelen, gewoon op een moment dat het jou het beste uitkomt.
Niet opstarten en terugvallen, maar vaardigheden opdoen waar je heel je leven gebruik van blijft maken. Dit zodat je resultaten blijvend zijn.

15+ jaar ervaring in de fitness
Sinds 2005 ben ik bezig met krachttraining en ben ik al meer dan 10 jaar werkzaam als (online) personal trainer.
Honderden mensen geholpen
Inmiddels heb ik honderden mensen geholpen met vetverlies, spieropbouw en een leefstijl die het leven verrijkt.
Wetenschappelijke vertaalslag
De adviezen die je krijgt zijn onderbouwd en makkelijk om toe te passen. Dit zodat je geen tijd en energie verspilt aan wat niet werkt en resultaat behaalt.
De maanden dat Victor me met mijn cut heeft begeleid, zijn me heel erg goed bevallen. Victor pakt alles op een wetenschappelijke en systematische manier aan. Dit was exact wat ik zocht. Het gaf mij direct veel vertrouwen in het coachingstraject. Ook vond ik het erg fijn dat ik alles kon vragen en hier goed onderbouwde reacties op kreeg. Mijn cut is nu klaar, en ik ben enorm tevreden met het resultaat! Ik ben van 88kg naar 77kg gegaan. En ik ben alleen maar sterker geworden! Daarnaast heb ik heel veel geleerd van dit traject en het was m’n geld meer dan waard. Nu gaat de volgende fase van start waarbij Victor mij begeleidt met mijn bulk! Ik zou Victor aan iedereen aanraden die zonder bullshit zijn droomfysiek wil behalen.

Sander van Strijp
Mijn programma’s
Online programma’s voor vetverlies en spieropbouw
Tijdens mijn online programma’s combineer ik jarenlange praktijkervaring, bewezen strategieën en de meest recente wetenschappelijke inzichten in een pragmatische aanpak.
Hiermee haal jij de resultaten waar je naar op zoek bent.
Afhankelijk van je doelen, heb je na een periode van 3 maanden de vaardigheden voor het leven opgedaan die nodig zijn om je doelen te behalen.
Persoonlijk. Toepasbaar. Onderbouwd.
Mijn e-books
Artikelen
- Vullend eten met weinig calorieën: mijn top 5!
 Vullend eten met weinig calorieën gaat jou op weg helpen om het afvallen makkelijker vol… Lees meer: Vullend eten met weinig calorieën: mijn top 5!
Vullend eten met weinig calorieën gaat jou op weg helpen om het afvallen makkelijker vol… Lees meer: Vullend eten met weinig calorieën: mijn top 5! - De 4 beste triceps oefeningen voor massieve triceps!
 Wat zijn de beste triceps oefeningen om je triceps te trainen? Door je triceps te… Lees meer: De 4 beste triceps oefeningen voor massieve triceps!
Wat zijn de beste triceps oefeningen om je triceps te trainen? Door je triceps te… Lees meer: De 4 beste triceps oefeningen voor massieve triceps! - Wat zijn de beste proteine repen (eiwitrepen)? Mijn top 3
 Proteine repen zijn erg populair. Ze zijn namelijk eiwitrijk, praktisch om mee te nemen en… Lees meer: Wat zijn de beste proteine repen (eiwitrepen)? Mijn top 3
Proteine repen zijn erg populair. Ze zijn namelijk eiwitrijk, praktisch om mee te nemen en… Lees meer: Wat zijn de beste proteine repen (eiwitrepen)? Mijn top 3

In 2023 heb ik de review awards
gewonnen bij coachscore.